
Пресс-центр / новости / Наука /
Происхождение, эволюция и разнообразие φ29-подобных фагов
Применение определения таксономического вида к описанию эволюционной истории биологических объектов с химерными геномами является сложной задачей. Ограниченный период существования видов и лабильность вирусных геномов могут сделать попытки реконструкции эволюционной истории практически бессмысленными. Процессы генетического обмена, по-видимому, особенно выражены у умеренных фагов, но генетические перестройки между эволюционно близкими группами фагов могут также затрагивать и φ29-подобные фаги. Тем не менее, можно попытаться построить эволюционные черты генов, кодирующих отдельные незаменимые белки и их стабильные комплексы.

Группа φ29-подобных фагов обладает выраженными уникальными особенностями в упаковке и репликации, которые являются взаимосвязанными процессами. Фаги этой группы имеют схожую архитектуру своих геномов и схожие вирионные белки (например, капсидные и портальные белки). Структурные взаимодействия между белками морфогенеза являются причиной коэволюцией этих белков и также могут быть одной из причин стабильности геномов φ29-подобных фагов. Поиски HMM и BLAST с использованием последовательностей основного капсидного белка и терминазы, проведенные в этом исследовании, позволили нам идентифицировать родственные фаги. Результаты показывают, что набор основных белков вириона (p6-p11) является отличительной чертой φ29-подобных фагов (рис. 1). Белки, составляющие репликационный комплекс и терминазу, также характерны для этих фагов.
Результаты исследования показывают, что эволюция φ29-подобных фагов сопровождалась перестройками генома, дупликацией и потерей генов, а также приобретением доменов, по-видимому, клеточного происхождения. Последнее относится даже к такому консервативному белку, как главный капсидный белок (major capsid protein, MCP). Интересно, что сравнение структур основного капсидного белка φ29 (исключая домен Ig-подобный, предположительно приобретенный путем горизонтального переноса) показало, что MCP φ29 больше похож на свой аналог из фагов Autographiviridae, чем на белки из других групп фагов Caudoviricetes (рис. 2). Это может быть аргументом в пользу эволюционного родства φ29-подобных и T7-подобных фагов. Однако непохожесть аминокислотных последовательностей MCP φ29-подобных фагов на последовательности MCP всех других вирусов Duplodnaviria (кроме фага KSY1), включая T7-подобные фаги, является убедительным подтверждением древнего происхождения φ29-подобных фагов.
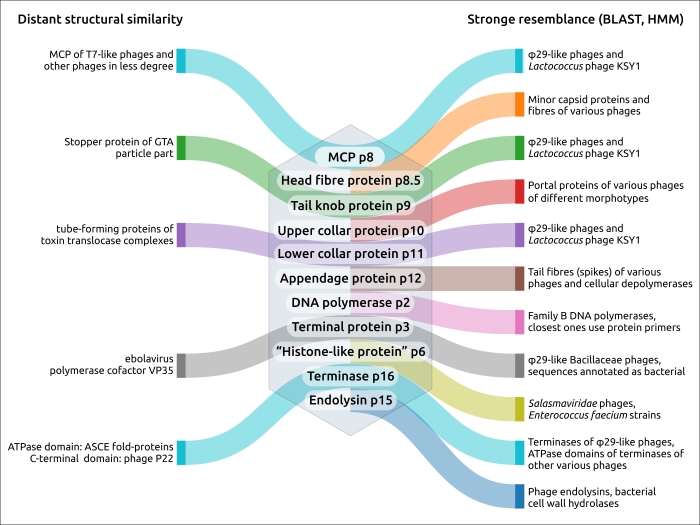
Рисунок 1. Диаграмма, показывающая близкое и отдаленное родство белков φ29, основанная на проведенных анализах и ранее опубликованных данных.
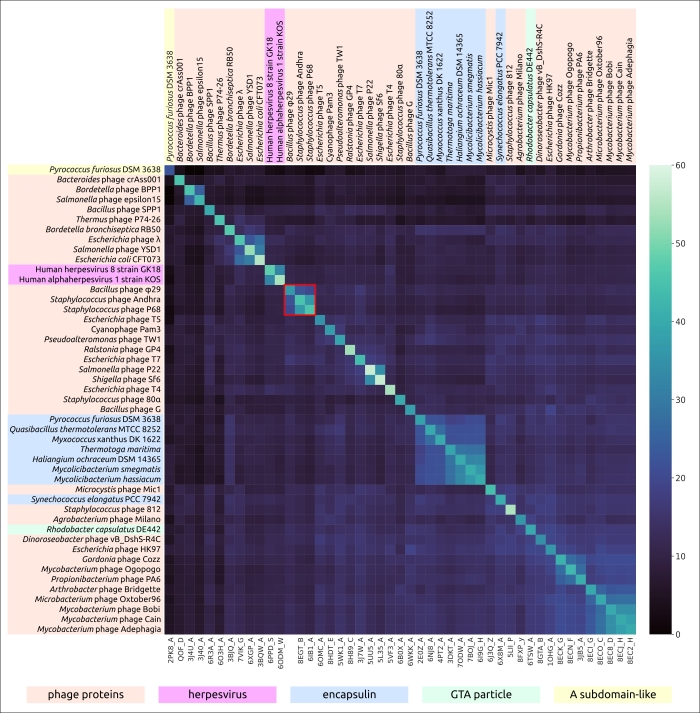
Рисунок 2. Тепловая карта, основанная на попарном структурном сходстве вирусных капсидных белков и основных белков оболочки инкапсулина, измеренном с помощью DALI Z-score. Кластер φ29-подобных фагов обведен красным.
Неожиданно поиск и использованием HMM показал, что необычный фаг Lactococcus KSY1 (Chopinvirus KSY1), который очень непохож на φ29 по размеру и составу генома, имеет φ29-подобную терминазу и белки вириона, включая отдаленные гомологи MCP φ29 и хвостовых белков. Возможно, φ29-подобные фаги являются всего лишь частью еще более крупной клады эволюционно связанных бактериофагов. По-видимому, фаг Lactococcus KSY1 приобрел гены адсорбционного аппарата от других фагов, инфицирующих Lactococcus или родственные бактерии, но источник генов, участвующих в транскрипции и репликации, требует дополнительных исследований. Поиск последовательности ясно указал на родство РНК-полимераз Chopinvirus, Autographiviridae и Schitoviridae, а филогенетический анализ показал, что РНК-полимеразы всех трех таксонов принадлежат к разным кладам. Интересно, сможет ли фаг Lactococcus KSY1 помочь найти связь между T7- и φ29-подобными фагами. Несмотря на наличие транскрипционного аппарата, напоминающего T7-подобные фаги, геном фага KSY1 не содержит ни T7-подобной ДНК-полимеразы, ни φ29-подобной ДНК-полимеразы. Согласно результатам филогенетического анализа, последняя связана с ДНК-полимеразой тективирусов.
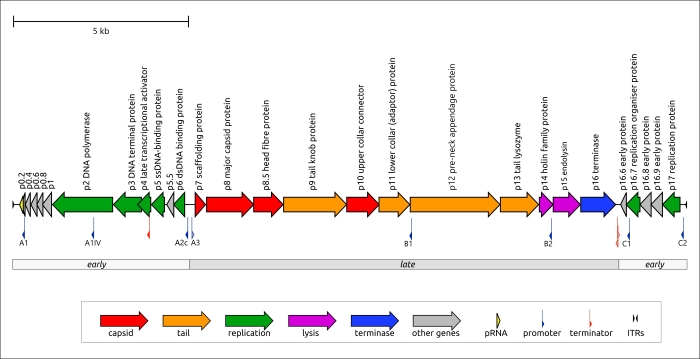
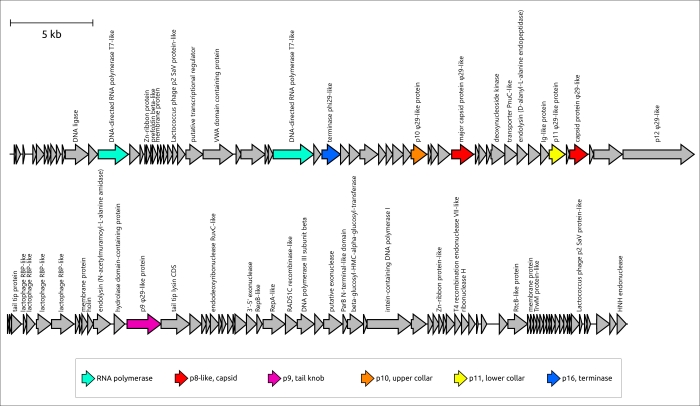
Рисунок 3. Генетическая карта фагов Bacillus φ29 и Lactococcus KSY1. Стрелки указывают направление транскрипции. Масштабная линейка указывает длину нуклеотидной последовательности. Функции генов указаны в подписях и легенде.
Хотя некоторые эволюционные родственники фагов с длинным хвостом известны, детали эволюции фагов с коротким хвостом изучены меньше. На самом деле, некоторые домены структурных белков фагов, особенно те, которые участвуют в адсорбции и деградации клеточной стенки, могут иметь сходство с белками или белковыми доменами клеточного происхождения, а также с белками вирусного происхождения, даже теми, которые закодированы в геномах фагов, принадлежащих к далеким вирусным группам, но эти случаи, по-видимому, являются результатом горизонтального переноса. Результаты исследования, которые указывают на сходство хвостовой трубки φ29, образованной белком p11, и хвостовых трубок комплексов токсин-транслоказы грамположительных бактерий, могли бы предложить частичное объяснение происхождения хвоста φ29-подобных фагов (рис. 4). Поиск схожих структур с использованием хвостовых белков φ29, за исключением рецептор-связывающих белков и портального белка, который является общим для всех хвостатых фагов и герпесвирусов и который можно считать частью капсида, не обнаружил схожих белков среди подовирусов, принадлежащих к хорошо изученным группам T7- (семейство Autographiviridae) и N4-подобных фагов (семейство Schitoviridae). В другой ранней публикации указывается, что аминоконцевой домен хвостовых белков φ29-подобного фага C1, инфицирующего Streptococcus, и, по-видимому, самого φ29, имеет укладку, обнаруженную в трубкообразующих белках фагов сифо- и миовирусов, что указывает на эволюционные связи между длиннохвостыми и короткохвостыми фагами. Подобная укладка была обнаружена в протеазах фагов и герпесвирусов, а также в белке-стопоре хвоста GTA Rhodobacter capsulatus, структурно напоминающем p11 согласно результатам DALI. Однако эта укладка, видимо, отсутствует в структуры белков другой подовирусной хвостовой трубки, а именно фага T7 (см., например, PDB #6R21). Возможно, морфологическая классификация подовирусов нуждается в некотором уточнении, поскольку их хвосты могут иметь разное происхождение и архитектуру.
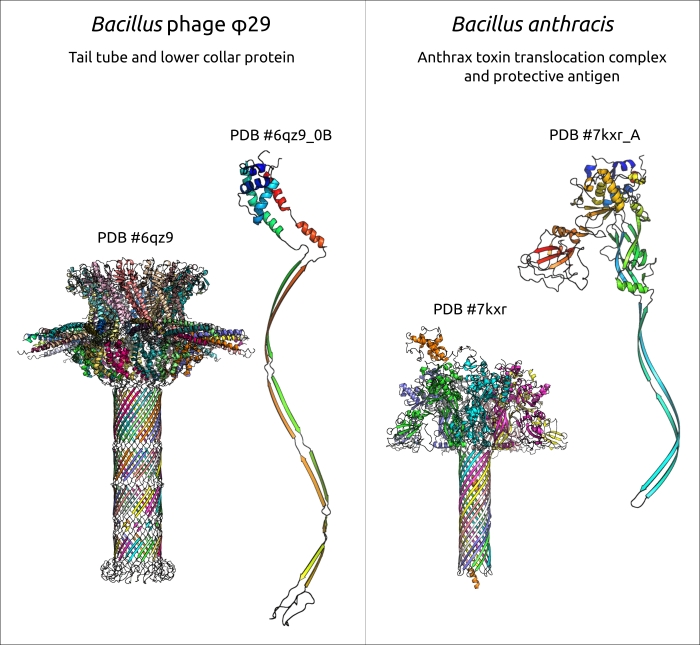
Рисунок 4. Хвостовая трубка фага φ29 и нижний воротниковый белок p11, а также комплекс транслокации сибиреязвенного токсина и защитный антигенный белок из Bacillus anthracis. В тройных комплексах разные цепи имеют разный цвет. Отдельные мономеры показаны с использованием радужной окраски с цветовым градиентом, где N-конец синий, а C-конец красный.
Эволюция φ29-подобных фагов, может характеризоваться как коэволюцией с хозяином, так и сменой хозяина, что можно предположить исходя из анализа и сравнения филогенетических деревьев, относящихся к последовательностям фагов и бактерий. φ29-подобных фагов способны инфицировать широкий спектр хозяев, а анализ метагеномных последовательностей показал, что, помимо бактерий порядков Bacillales, Clostridia, Enterobacterales и Pseudomonadales, эти фаги могут инфицировать представителей порядка Bacteroidales. Возможно, фаг Bacteriophage sp. изолят 2692_55609, представляющий кластер c30, является самым ранним дивергировавшим фагом, подобным φ29 (среди проанализированных представителей), который инфицирует грамотрицательные бактерии. Филогенетический анализ MCP и ДНК-полимеразы показывает, что этот фаг связан с фагом Amedibacillus AD70P2. Бактерии Amedibacillus филогенетически далеки от грамотрицательных бактерий. Происхождение и эволюция фагов, подобных φ29, инфицирующих грамотрицательные бактерии, вероятно, связаны с переключением хозяина, за которым следует модификация некоторых белков. В частности, фаги, инфицирующие грамотрицательные бактерии, имеют укороченный lower collar protein (и хвостовую трубку), что может быть связано с более тонкой клеточной оболочкой бактериального хозяина. Кроме того, недавняя публикация о фаге Acinetobacter Phanie и результаты этой работы предполагают особый состав хвостового выступа (tail knob) у фагов, инфицирующих грамотрицательные бактерии (рис. 5). “Tail knob” φ29-подобных фагов, инфицирующих грамотрицательные бактерии, по-видимому, представляет собой гексамер, каждая единица которого состоит из двух белков. Один из них расположен ближе к головке фага и соответствует N-концевой части белка tail knob φ29 (p9), а второй дистальный белок аналогичен карбоксиконцевой части p9. Расщепление гена, кодирующего белок хвостового выступа, вероятно, произошло у предка φ29-подобных фагов, инфицирующих грамотрицательные бактерии, вследствие генетического обмена и привело к вставке фрагмента, кодирующего Ig-подобные домены. Ранее было показано, что Ig-подобные домены могут участвовать во взаимодействии с углеводами на поверхности бактериальной клетки и участвовать в формировании хвостовых придатков и оболочек фага. Результаты структурных предсказаний предполагают, что Ig-подобные домены расположены латеральнее хвостовой трубки. В частности, модель AF белкового комплекса хвостового выступа фага Pectobacterium DU_PP_III, предсказанная с приличным качеством (оценка ipTM 0,69, оценка pTM 0,70) (рис. 6), изображает Ig-подобные домены как напоминающие короткие хвостовые волокна, хотя и с архитектурой, со структурной архитектурой, принципиально отличной от обычной тримерной архитектуры. Фаг Acinetobacter Phanie содержит шесть таких доменов в белке gp11. Они напоминают Ig-подобный некаталитический домен L,D-транспептидазы из Mycobacterium tuberculosis; гипотетически эти домены способны проявлять свойства связывания углеводов. В предыдущей работе юыло экспериментально продемонстрировано образование химерных вирионов фага Phanie и хелперного миовируса Acinetobacter phage Aci01-1. Согласно данным электронной микроскопии трубка хвоста миовируса прикреплена дистально к хвосту фага Phanie. В одной из предыдущих работ было высказано предположение, что дополнительные Ig-подобные домены белка оболочки хвоста миовируса могут способствовать повышению стабильности оболочки за счет белок-белковых взаимодействий с участием дополнительных доменов. Вероятно, эти домены играют важную роль в образовании химерных вирионов.
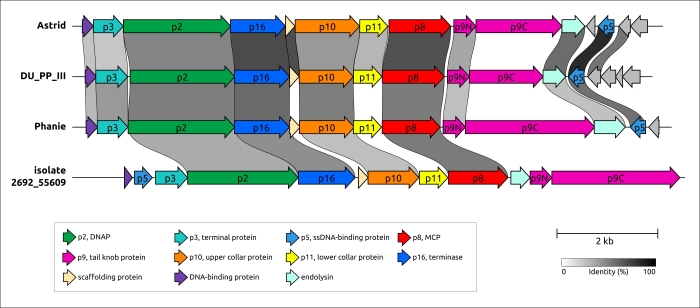
Рисунок 5. Генетическая карта фагов, принадлежащих кластерам 23 и 30. Названия фагов следующие: Astrid, Salmonella phages Astrid; DU_PP_III, Pectobacterium phage DU_PP_III; Phanie, Acinetobacter phage Phanie; isolate 2692_55609, Bacteriophage sp. isolate 2692_55609. Аннотации, основанные на результатах поиска HMM, предсказаниях AF и ранее опубликованном геномном анализе фага Phanie, показаны в подписях. Стрелки указывают направление транскрипции. Градинетная полоска указывает уровень идентичности между генами в соответствии с черно-белым градиентом, легенда для которого показана в нижней части рисунка. Количество кластеров и название последовательностей показаны слева. Гены, кодирующие белки, подобные φ29 (p2, p5, p6, p8–p11, p16) и другие белки, окрашены в соответствии с легендой.
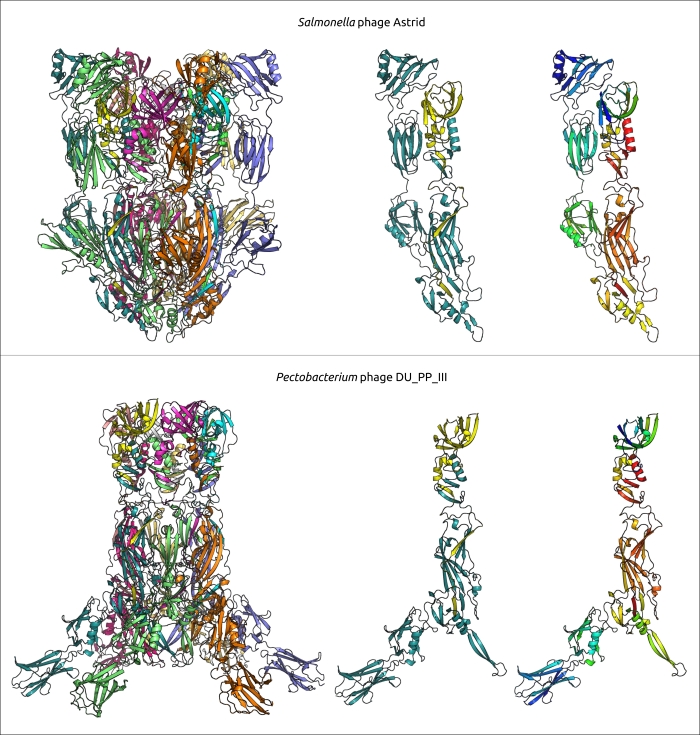
Рисунок 6. Ленточная диаграмма белковых комплексов, смоделированных AlphaFold3, предположительно соответствующих хвостовым выступам фагов Salmonella Astrid и фага Pectobacterium DU_PP_III. Левые изображения показывают полноразмерную модель, состоящую из 12 мономеров (каждый мономер имеет свой цвет). Центральное изображение показывает два самых близких различных мономера, белок, похожий на N-концевую часть белка хвостового выступа φ29, окрашен в желтый цвет, а белок, похожий на C-концевую часть, окрашен в голубой цвет. Правые изображения показывают те же мономеры, что и центральные, с использованием радужного цветового градиента, где N-концевой конец каждого мономера синий, а C-конец красный.
Подводя итог вышесказанному, группа фагов, подобных φ29, явно отличается от других групп фагов. У них много общего, хотя и есть различия в геномах, бактериальных хозяевах и механизмах заражения. Поиск вирусов, похожих на фаг φ29 и родственные, не смог найти многих членов этой группы, поскольку он использовал HMM-методы на ограниченной базе данных. Однако поиск выявил более 2000 последовательностей, что указывает на большую распространенность этих вирусов. Распространенность φ29-подобных фагов, комплекс уникальных особенностей, включая своеобразные морфологию и процессы репликации, и их характерное происхождение, включая раннюю дивергенцию консервативных фаговых белков, предполагают возможность выделения этой группы в отдельный таксон. Возможно, ее следует отнести к отряду или таксону более высокого ранга.
Работа опубликована в журнале International Journal of Molecular Sciences.
11 октября 2024 года